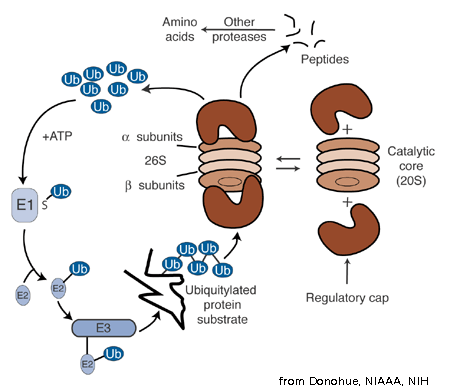
需要被蛋白酶体降解的蛋白质会先被连接上泛素作为标记,即蛋白质上的一个赖氨酸与泛素之间形成共价连接。这一过程是一个三酶级联反应,即需要有由三个酶催化的一系列反应的发生,整个过程被称为泛素化信号通路。在第一步反应中,泛素活化酶(又被称为E1)水解ATP并将一个泛素分子腺苷酸化。接着,泛素被转移到E1的活性中心的半胱氨酸残基上,并伴随着第二个泛素分子的腺苷酸化。 被腺苷酸化的泛素分子接着被转移到第二个酶,泛素交联酶(E2)的半胱氨酸残基上。最后,高度保守的泛素连接酶(E3)家族中的一员(根据底物蛋白质的不同而不同)识别特定的需要被泛素化的靶蛋白,并催化泛素分子从E2上转移到靶蛋白上。靶蛋白在被蛋白酶体识别之前,必须被标记上至少四个泛素单体分子(以多泛素链的形式)。 因此,是E3使得这一系统具有了底物特异性。 E1、E2和E3蛋白的数量依赖于生物体和细胞类型,人体中就存在大量不同的E3蛋白,这说明泛素-蛋白酶体系统可以作用于数量巨大的靶蛋白。
多泛素化后的蛋白质是如何被蛋白酶体所识别的,还没有完全弄清。泛素受体蛋白的N末端具有一个类泛素结构域,以及一至多个泛素结合结构域。类泛素结构域可以被19S调节颗粒所识别,而泛素结合结构域可以通过形成三螺旋束来结合泛素。这些受体蛋白可能能够结合多泛素化的蛋白质并将其携带到蛋白酶体,而关于这种结合的特异性和调控机制还不清楚。 但最近有研究者发现,调节颗粒上的亚基Rpn13可以发挥泛素受体的功能。
泛素蛋白自身由76个残基所组成,以“泛素”为名是因为它在生物体中广泛存在:具有高度保守的序列并且存在于所有已知的真核生物体中。真核生物中编码泛素的基因以串联重复(tandem repeat)的方式排列,这可能是因为大量转录的需要,为细胞生产足够缺樱滚多的泛素。有人提出泛素是目前发现的进化速度最慢的蛋白质。 泛素化信号通路。其中,“Ub”表示泛素。
泛素化后的蛋白质(以下称为底物蛋白)被19S调节颗粒所识别,这一过程是一个ATP依赖的结合过程。 然后,底物蛋白必须进入20S核心颗粒的内部孔道,以便与位于其中的水解活性位点接触。由于20S颗粒的孔道相对狭窄,而且两端由α环中亚基的N末端控制开关,所以底物蛋白在进入核心颗粒之前必须至少部分去折叠。将去折叠的蛋白质传递进入核心颗粒的过程被称为“移位”(translocation),而移位必须发生在去泛素化之后。 但目前对于底物蛋白的去泛素化和去折叠机制还不了解。 在整个降解反应过程中,那一步是限速步取决于底物蛋白的类别;对于一些蛋白质,去折叠过程是限速步,而对于另一些蛋白质,可能是去泛素化为限速因子。 至于哪些底物蛋白在移位之前必须去折叠,还未有结论,而牢固的三级结构和一些特殊的非局部相互作用,如二硫键,能够抑制降解。
由α亚基所形成的“门”可以阻止长于四个残基的多肽进入20S颗粒的内部。在识别步骤开始前结合上的ATP分子在移位发生前被水解,而对于水解产生的能量是用于蛋白质去折叠 还是“门”的打开 还有争议。26S蛋白酶体在存在无法水解的ATP类似物(即无法获得水解产生的能量)的情况下,依然可以降解去折叠的蛋白质,但却无法降解折叠的蛋白质;这一结果说明ATP水解所产生的能量至少部分被用于蛋白质去折叠。 在19S帽子处于伏余ATP结合状态时,去折叠的底物蛋白可以由促进扩散作用,传递通过开启的“门”。
球蛋白去折叠的机制是基本类似的,但在一定程度上也取决于蛋白质的氨基酸序列。研究者发现含有较长的甘氨酸或丙氨酸序列可以抑制去折叠,从而降低蛋白酶体的降解效率;其结果是生成含有部分去折叠蛋白质的混合物,这可能是由于ATP水解和去折叠步骤之间的脱节所导致的。 自然界中的一些蛋白质也有这样的甘氨酸-丙氨酸重复序列存在,如蚕丝中的丝心蛋白(fibroin);值得一提的是,特定的人类疱疹病毒基因的表达产物也含有这样的序列,通过抑制蛋白酶体的作用,阻止了抗原呈递到主要组织相容性复合体上,从而有助于病毒的繁殖。
20S核心颗粒的一个剖面图,显示了活性位点的位置。其中,α亚基用绿颂高色的球来表示,β亚基的蛋白骨架显示为飘带,并且不同的多肽链用不同的颜色表示。小的粉色球表示每个亚基的活性位点中苏氨酸残基的位置。淡蓝色的化学结构为结合在活性位点上的抑制剂硼替佐米(bortezomib)。 蛋白质的降解由20S核心颗粒中的β亚基进行,其机制被认为是苏氨酸依赖的亲核攻击。这一机制可能需要有一个结合的水分子参与活性的苏氨酸上羟基的去质子化。降解发生在核心颗粒中间的两个β环内的孔道里,一般不生成部分降解的产物,而是将底物蛋白完全降解为长度一定的肽段;肽段的长度一般为7-9个残基,但根据生物体和底物蛋白的不同,长度范围可以从4-25个残基不等。决定分解产物中肽段长度的机制,目前还没有完全弄清。 虽然具有催化活性的三个β亚基具有共同的降解机制,但它们对于底物的特异性却略有不同,分别为类胰凝乳蛋白酶型、类胰蛋白酶型和肽谷氨酰基肽水解型。这种对于底物特异性的差异是来自于靠近活性位点的局部残基与底物之间的相互作用的不同。每一个具有催化活性的β亚基也都含有一个降解所必需的保守的赖氨酸。
虽然蛋白酶体通常生成非常短的降解片断,但在一些情况下,这些降解产物自身是具有生物学活性的功能分子。特定的转录因子,包括哺乳动物的NF-κB复合物中的一个组分,合成后是以无活性的前体分子存在,在经过泛素化和蛋白酶降解后,才转变为活性分子。这种降解需要蛋白酶体剪切蛋白质的中间部分,而不是通常情况下的从蛋白质的一端开始的剪切。有人提出,需要被剪切的中间部分为一个长的loop,位于蛋白表面,从而可以作为蛋白酶体的底物进入其内部孔道,而蛋白质的其他部分依然在孔道外,并不会被降解。 在酵母蛋白中也发现了类似的现象;这种选择性降解被称为“受调控的泛素-蛋白酶体依赖的剪切”(regulated ubiquitin/proteasome dependent processing)。 虽然大多数的蛋白酶体的底物必须在降解之前被泛素化,但仍然有一些例外的情况,尤其是在蛋白酶体参与蛋白质的翻译后处理过程中。一个主要的例子是蛋白酶体通过将p105蛋白剪切为p50蛋白来激活NF-κB。 一些由于存在无结构区域(参见intrinsically unstructured proteins)而被推测具有不稳定性的蛋白质也可以通过非泛素依赖的途径被降解。鸟氨酸脱羧酶是最著名的非泛素依赖途径中蛋白酶体的底物。 对于关键的细胞周期调控因子,如p53蛋白的非泛素依赖的降解机制已经有报道,虽然p53蛋白也可以通过泛素依赖的途径被降解。 此外,在一定的细胞应激条件下,结构不正常、错误折叠或者过度氧化的蛋白质也都会进入非泛素依赖的和非19S颗粒依赖的降解途径。